Gross pathology:
With the organ block positioned such that the posterior surface is facing upward, the adrenals can easily be dissected by incising and lifting up the posterior diaphragm. At this point I take a sample of the diaphragm for histology, trying to get the muscle fibers in cross section. Removing most of the diaphragm facilitates the dissection. The adrenals should be immediately seen beneath the diaphragm and are superior and partly deeper to the kidneys. They can be weighed, usually as combined weight if symmetrical. Standard histological samples are taken transversely through the center of each gland. If the glands are from very early gestation, they may be submitted uncut. For fat stains, a separate section can be frozen. Fat stains an also be done on frozen sections of formalin fixed tissue prior to paraffin embedding.
A study of adrenal measurements and weights in fetal autopsies from 15 to 27 weeks of gestation showed a linear increase in adrenal weight from 0.2-1.5 g[1]. Most of the fetuses were from spontaneous losses and a few from medically indicated abortions, with the main analysis confined to those with little autolysis. Weights were decreased in fetuses with severe autolysis. The spread of values was wide, and although some underlying conditions were noted especially in reviewing cases with cytomegaly or hyperplasia too little information was given to evaluate the effects of disease. Lipid appeared to decrease with advancing gestation, and some fat stains were performed but the illustrations are with Hematein Phloxin Safran in which the lipid appears to be clear vacuoles.
A study of adrenal weights from 182 elective abortions from 6 to 17 weeks of gestation based on foot length found a rapid increase after 12 weeks of gestation[2]. The weight can still be compared to the standard adrenal brain ratios reported from the Perinatal Collaborative Project[3]
The major gross pathologic findings are hypoplasia and lipid accumulation. The adrenals in the perinatal period are relatively larger compared to the kidney than they are later in childhood, and in adults. This is due to the large fetal cortex that produces the estrogen precursor DHEA-S and progesterones in utero, but involutes in the weeks to months after birth. Except with stress as expressed by increased ACTH, the adrenal does not accumulate lipid, and thus does not have the typical yellow color seen in pediatric and adult adrenals. However, in some conditions, lipid accumulates in the fetal adrenal and presents grossly as a distinct yellow color (see microscopic below). (Fig 1, 2)


Other observations that may be seen are asymmetry of the two adrenals due to hemorrhage or tumor. If kidneys are not present there will be a distortion of shape with the adrenal extending into the area of the missing kidneys, an anatomic distortion that initially fooled early fetal ultrasonographers into identifying non-existent kidneys.
A standardized dictation for normal: The adrenals are in their normal location, and are approximately one quarter the size of the kidney and are pink in color.
Gross abnormalities to note:
- Small or large size in relation to the kidney: Small adrenals occur with interruption of the pituitary adrenal axis as in holoprosencephaly. They are also abnormally small in triploidy, although the mechanism is not fully determined.
- Yellow color reflecting lipid: This occurs with stress and is associated with a thick permanent cortex identified microscopically. Lipid accumulation also occurs in lipid storage disease such as glutaric aciduria Type 2.
- Abnormal shape of the adrenals: This is usually the consequence of an absent kidney with adrenal appearing larger in area and flattened.
- Congestion and or hemorrhage: While generalized congestion can occur with heart failure, prominent adrenal hemorrhage expanding or disrupting the gland if often a finding in shock, particularly septic shock.
- Small nodules of adrenal tissue just outside the capsule.
- Absent adrenals. The para-aortic area should be sampled to look for nodules of cortex. Paraganglionic nodules may be present with the larger organ of Zuckerkandl in this area.
- Tumors and cysts including congenital neuroblastoma
The microscopic findings:
The normal gland: The adrenal cortex is composed of a narrow definitive zone of cortical cells with sparse clear cytoplasm and a thick fetal cortical zone of plump eosinophilic cells lining sinusoids. A 2002 review recapitulates the well-known development of the primate fetal adrenal cortex which produces DHEAS but not cortisol and uses progesterone as a substrate which may include placental progesterone[4]. The placenta converts DHEAS to estrogen (estriol) which has a positive feedback loop on steroidogenesis from cholesterol. ACTH stimulates fetal DHEAS production. The authors point out the unusual relationship of placental corticotrophin releasing factor which is stimulated by corticosteroids creating a positive feedback loop, and the CRF releasing factor receptors in the fetal adrenal are coupled to phospholipase and phosphoinositol rather than adenylate cyclase. DHEA production starts around 8 weeks of gestation, but the adrenal develops independently of ACTH until 15 weeks of gestation as evidenced by adrenal hypotrophy in anencephaly. The authors also note that the P450 aromatase mutations affecting estrogens, and estrogen receptor mutations do not result in abnormal parturition. There is a thin definitive zone cortex which produces aldosterone. The fetal cortex will involute and the definitive cortex will develop into the postnatal steroid producing organ. By mid-gestation the inner fetal zone produces DHEA, and a mid-zone produces cortisol. The mid region or transitional zone is not morphologically distinct from the inner fetal zone by the usual H&E pathology. Both regions are stimulated by ACTH to increase hormone output. Growth after 15 weeks of gestation is also dependent on ACTH stimulation of paracrine growth factors. The negative feedback of cortisol on ACTH production is present. Placental CRF (corticotrophin releasing factor) may also play a regulatory role either on ACTH or directly on the adrenal cells.
Medulla and catecholamines:
In the newborn it is a thin layer of immature immunoblasts, less than 1% of volume[5]. In younger gestation fetuses usually below 16 weeks gestation there will be nodules of neuroblasts that resemble the small blue cells of neuroblastoma. Congenital neuroblastoma may be found in the adrenal.
A 1983 review of fetal catecholamine studies found several well established observations[6]. Catecholamines, primarily norepinephrine, are secreted from paraganglia in the fetus. They are secreted in response to fetal hypoxia, and result in a redistribution of blood favoring the heart, brain, adrenal, and placental blood flows. Endogenous catecholamines do not transfer across the placenta. Catecholamines may increase fetal production of prostaglandins, but there was no firm evidence of their role in initiating labor.
Autolysis:
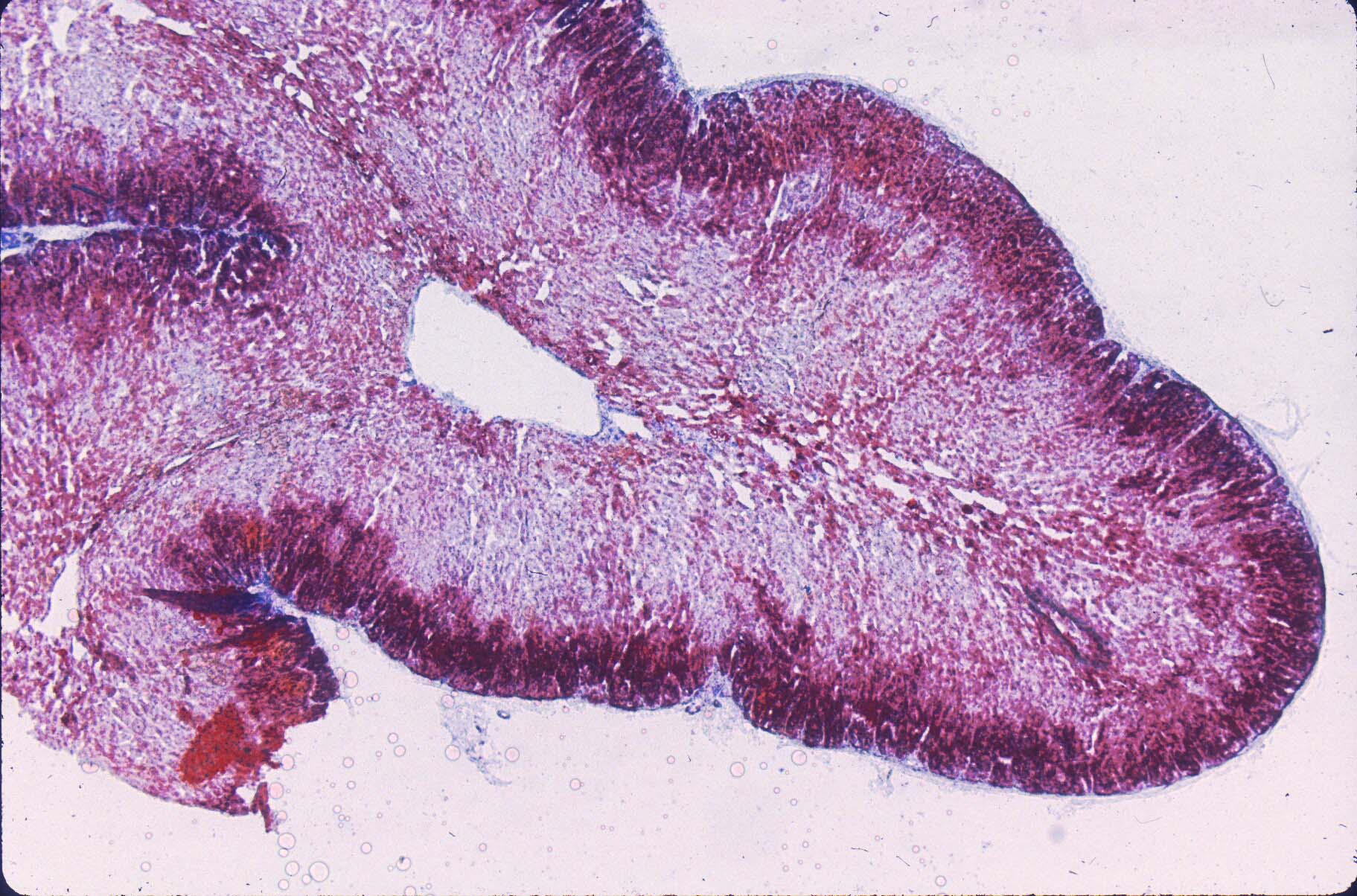
In the Genest grading, complete loss of basophilia occurs after 1 week, but the degree of autolysis is often very marked even if not complete. This may obscure the evaluation on H&E slides of lipid accumulation, but the identification of loss of nuclear basophilila will still be discernible. (Fig 3)
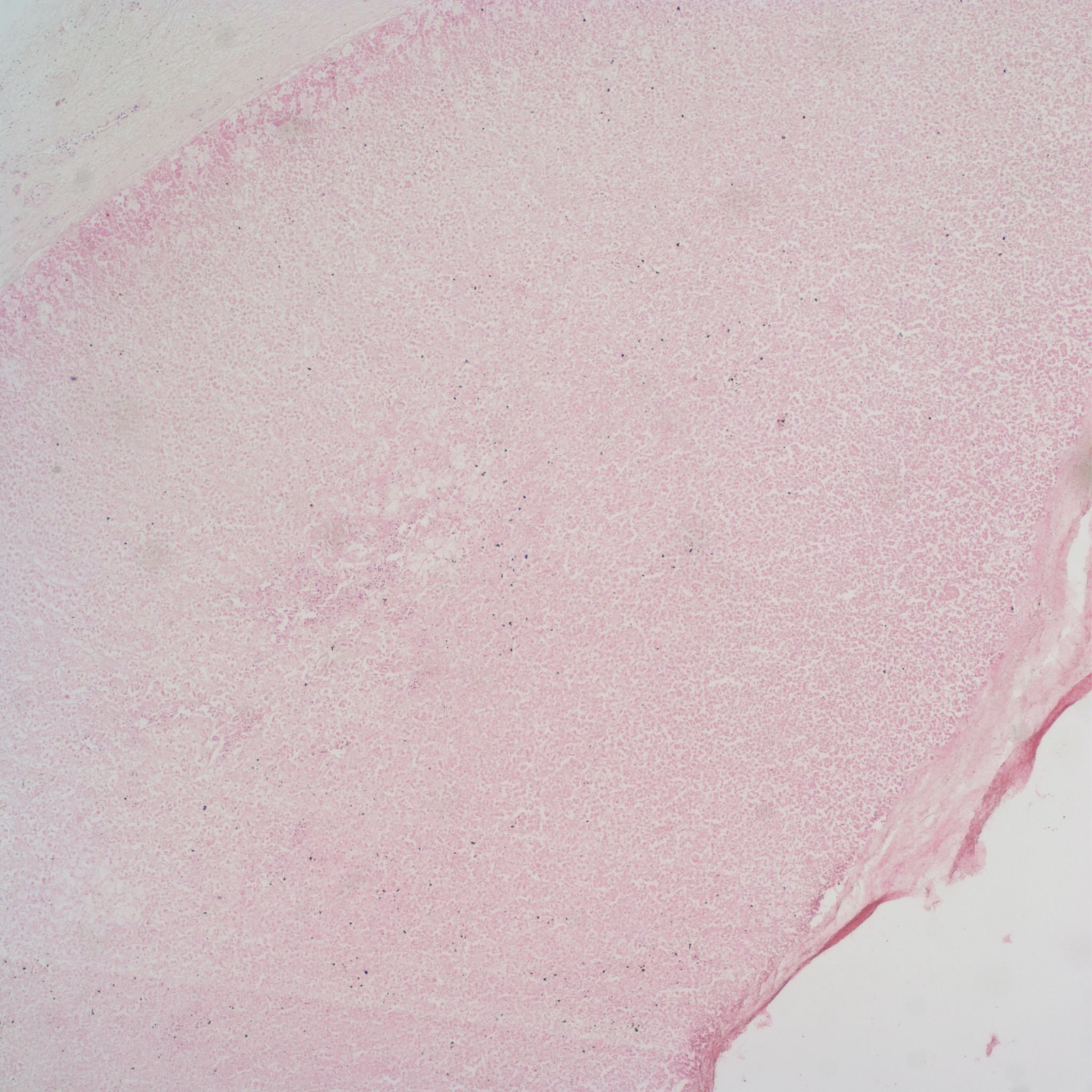
Lipid accumulation in the fetal cortex:
Microscopically the lipid is seen as clear vacuolization of fetal adrenal cells (Fig 4).

Without a fat stain of tissue that has not been processed through graded solvents to remove fat, the vacuolization of the cells could be from swelling or autolysis. Gross confirmation of the lipid or a fat stain aid the diagnosis increase in lipid (Fig 5).
An autopsy study of formalin fixed fat stained adrenal glands in 40 infants 22-41 weeks of gestation and with 2 days to 10 weeks of postmortem retention[7] defined three patterns of fat distribution. Stage I had no lipid. Stage II had a zone of little fat between the permanent and fetal cortex. Stage three had continuous zone of fat throughout. The fifteen stage 1 cases correlated with normal placentas and histories of asphyxia or trauma. The 8 infants with stage 2 correlated with hydrops including syphilis and Rh alloimmunization. The 17 infants with stage 3 had circulatory disorders of the placenta (very broadly defined but had to involve 25% of the placenta and was often associated with toxemia). Typically, there are yellow adrenals in infants with fetal hydrops. That there could be a difference in fat patterns between preeclampsia and hydrops is plausible.
Both the permanent and the fetal cortex respond to ACTH by increased production, but the finding of fat in the cells is not necessarily evidence of increased steroid hormone production. A study comparing the umbilical cord levels of DHEA sulfate and of lipoprotein-cholesterol (fetal adrenal substrate for steroidogenesis) found the lowest levels of the first and highest of the latter in infants from 37 mothers with severe preeclampsia compared to129 infants from uncomplicated pregnancies[8]. In umbilical cord plasma comparing normal to mothers with severe pre-eclampsia, and to anencephalic infants, there is a direct inverse relationship between low density lipoprotein cholesterol and DHEA[9]. A clinical study of ACTH stimulation in preterm growth retarded neonates demonstrated a decreased cortisol response compared to gestational matched controls[10]. The authors suggest that this persistent blunted response to ACTH may have a role in neonatal morbidity and mortality, and possibly play a role in the long-term problems of growth retarded premature infants. This evidence suggests that DHEA is being suppressed in pre-eclampsia and that the lipid accumulation in the adrenal is because of cholesterol back up. There is less information on hormone function in hydrops. However, both hydrops and severe utero-placental ischemia are associated with thymic involution that experimentally is triggered by cortisol, suggesting that function of the permanent cortex is not suppressed but in fact is increased by ACTH stimulated by either hypothalamic or placental CRF. The difference in the lipid accumulation recorded as stages II and III in the above study could simply be a matter of degree. Whatever, the physiologic details adrenal lipid is an indicator of stress that is associated with more or less chronic conditions, that on occasion may be a useful clue about intrauterine events.
There is an exception to adrenal lipid being an indicator of stress and that is lipid accumulation in metabolic disease such in mucolipidosis II or in glutaric acidemia Type II in which there is fat accumulation in multiple organs of which the one is the adrenal.
Compact and cystic areas in the definitive adrenal cortex;
These related lesions were evaluated in 75 stillborn infants by comparing them to thymic involution, abruption, and intrathoracic petechiae[11]. These lesions often occurred in the same sample and their presence correlated with a higher degree of thymic atrophy. The latter is a surrogate for the effect of cortisol on accelerated thymic involution. These definitive cortical lesions are likely a marker for prolonged fetal ACTH stimulation. The absence of the lesion correlated with acute asphyxia.
Adrenal Cytomegaly:
This is a poorly understood phenomenon in which individual cells in the fetal zone adrenal show a two-fold enlargement with large nuclei. The cells may be in one or more focal clusters or diffuse. In the pancreas, cytomegaly with large polyploid nuclei is associated with increased intrauterine hormone production, but in the adrenal the association of cytomegaly and increased DHEA is at best tenuous. Where it has been reported, the maternal level of estriol has been low in cases with adrenal cytomegaly[12]. Estriol is a fetal liver product that uses DHEA as a substrate that was measured in the past as a marker of fetal well-being. Series of autopsy cases (stillborn and neonatal), including an unpublished one in from Cincinnati Children’s Hospital of 32 cases from 1954 to 1976, found an excess of infants with preeclampsia, Rh alloimmunization, and twins which are similar associations with adrenal lipid accumulation[12, 13]. More recently adrenal cytomegaly has been reported to be frequent in hydrops from Bart’s hemoglobin showing that the immunological component of alloimmunization is not integral to the association of cytomegaly with anemia[14].
. Direct studies of cytomegalic cells by flow cytometry and by immuno-histochemistry and morphometry, support the polyploid origin of the large cells[15-17]. Neither normal nor cytomegalic fetal zone adrenal cells showed proliferation including one case with flow cytometry that showed low S phase cells. (Definitive zone adrenal cells did show numerous proliferative cells.) Of 9 nine fetal adrenals stained for the proliferation marker Ki-67, one of nine showed proliferation of fetal zone cells That infant had received antenatal steroids[17]. One hypothesis suggested to account for the findings is that the polyploid adrenal cells were initially hyperstimulated and then burnt out.
Adrenal cytomegaly may be associated with specific entities such a congenital adrenal hypoplasia, cytomegalic type and with Beckwith Wiedemann syndrome. One study highlighted the high number of infants with various malformations with adrenal cytomegaly which may reflect an autopsy bias rather than an association[12]. In older age cases of adrenal virilism and of Beckwith Wideman syndrome, the hyperfunctioning adrenal shows adrenal cytomegaly, but this is not fetal DHEA producing tissue[18]. It is important to distinguish adrenal cytomegaly from cytomegalovirus infection in the adrenal which produces giant cells in the fetal adrenal that have large nuclear inclusions
Adrenal cytomegaly is found in perinatal autopsies without other evidence of stress. Endocrine cells may become polyploid in response to a stimulus[16, 17]. These polyploid cells are thought to no longer be in the cell cycle. A reasonable hypothesis is that adrenal cytomegalic cells are the residua of an earlier intrauterine stimulus to the fetal adrenal[15]. Other possible mechanisms are that the cells are a spontaneous abnormal clone, or are a reaction to a focal stimulus such as virus. Adrenal cytomegaly is only identified postmortem, and therefore has no known clinical correlation if not associated with a genetic disease. Adrenal cytomegaly may be evidence of increased fetal ACTH stimulation in fetal hydrops or maternal preeclampsia, but there is insufficient evidence that the lesion is always a marker of ACTH stimulation. Theoretically, the polyploid cells could be a marker of earlier transient fetal stress, but there is currently no evidence for that hypothesis. Finding adrenal cytomegaly should alert the pathologist to exclude Beckwith Wiedemann syndrome.
Viral infection:
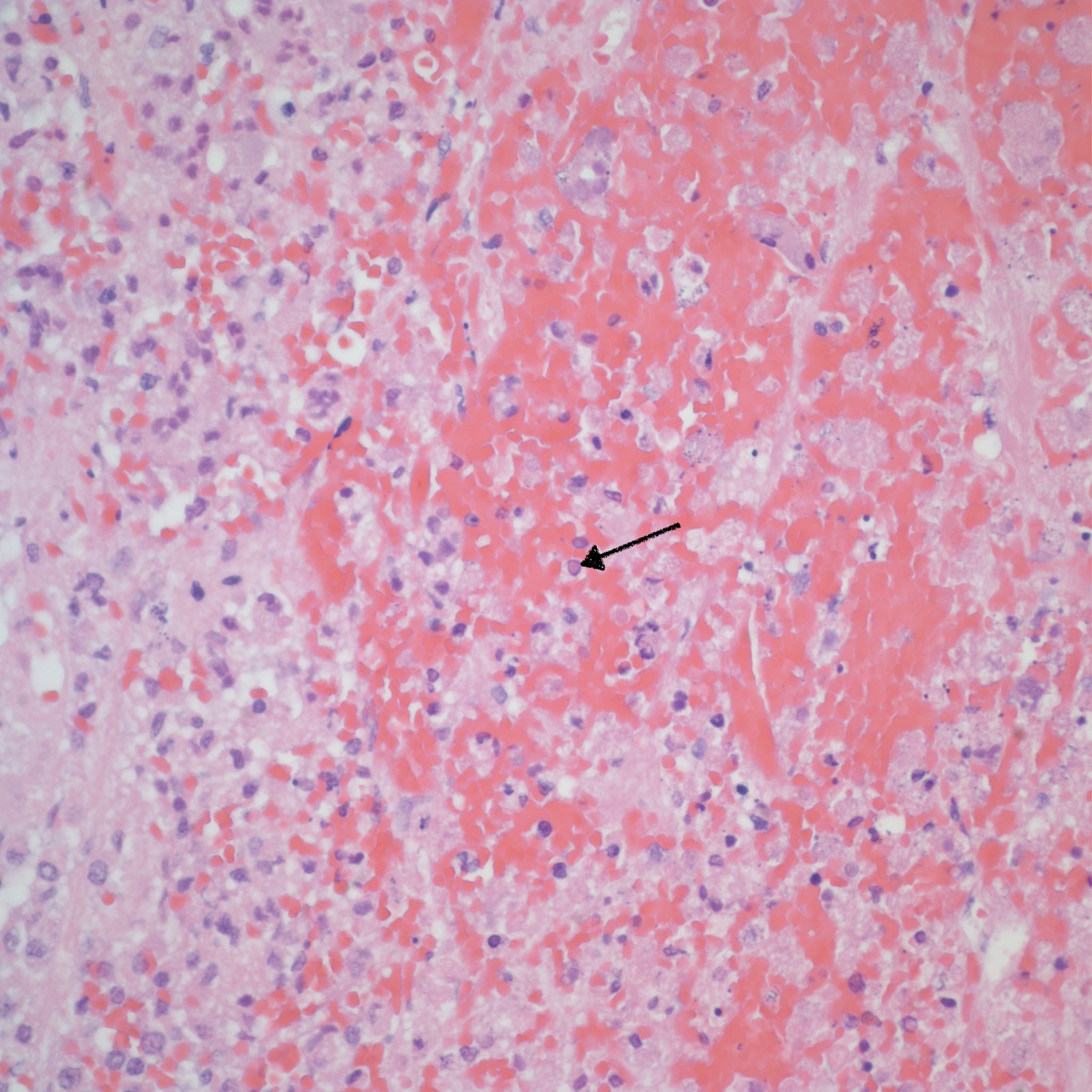
The adrenal gland is a frequent target in congenital herpes simplex infection and will demonstrate focal areas of cellular necrosis (Fig 6).

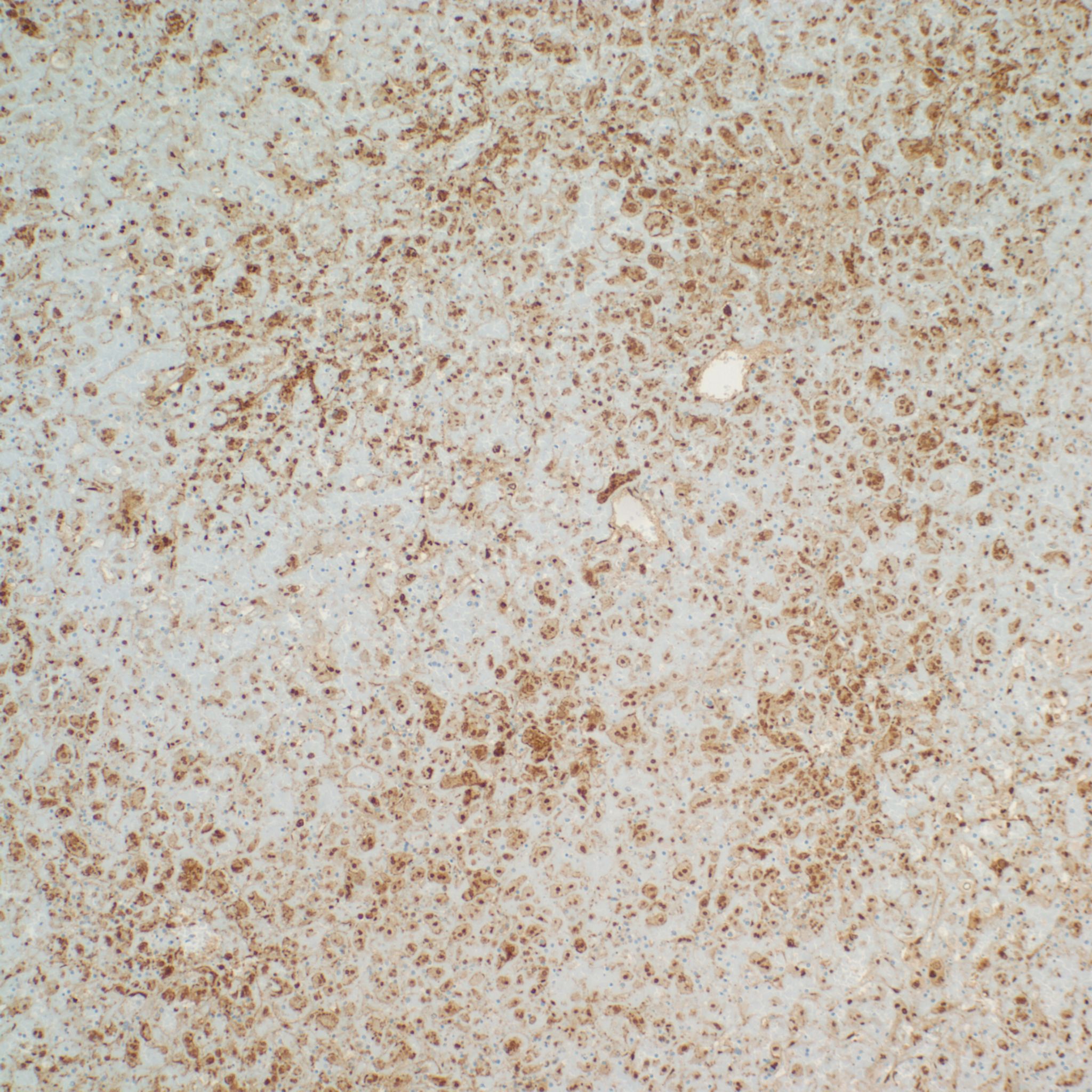
Usually at the periphery, herpetic inclusions are evidence, which can be confirmed with specific HSV 1, 2 immunostaining even in autolytic tissue (fig 7-8). It may infrequently be show viral inclusions in CMV infection.
Small adrenals:
Small adrenals in the fetus are usually due to relatively decreased development of the fetal cortex. The fetal adrenal cortex relies on ACTH stimulation. In anencephaly that is sufficiently complete to eliminate the hypothalamic-pituitary axis, the adrenal are small [19, 20]. There is a syndrome of familial ACTH deficiency which if not diagnosed can result in death in the first day of life and will demonstrate small adrenals[21, 22]. The mechanism of small adrenals is less clear in triploid fetuses, but it is a common finding[23-26]. The relative loss of fetal adrenal zone appears less than in anencephaly.
Large adrenals:
Symmetrical enlargement may be evidence of congenital adrenal hyperplasia which is not usually a cause of fetal death[27]. Asymmetrical enlargement of the gland is more likely to be due to hemorrhage, but tumor including congenital neuroblastoma may be present. There is a pseudo enlargement of the adrenal gland which is due to expansion of the gland caudally in cases of absent kidneys. The gland has greater areas but is flattened.
Hemorrhage, congestion, thrombi and infarctions:
Medical students are likely to be aware of adrenal hemorrhage as a manifestation of septic shock with the Waterhouse Friderichsen hemorrhages with meningococcemia. In the fetus and newborn, the first manifestation of shock is likely to be either congestion/hemorrhage or fibrin thrombi. The fibrin thrombi were documented as a neonatal response to shock in infants with hypoplastic left heart syndrome [28] (Fig 9).

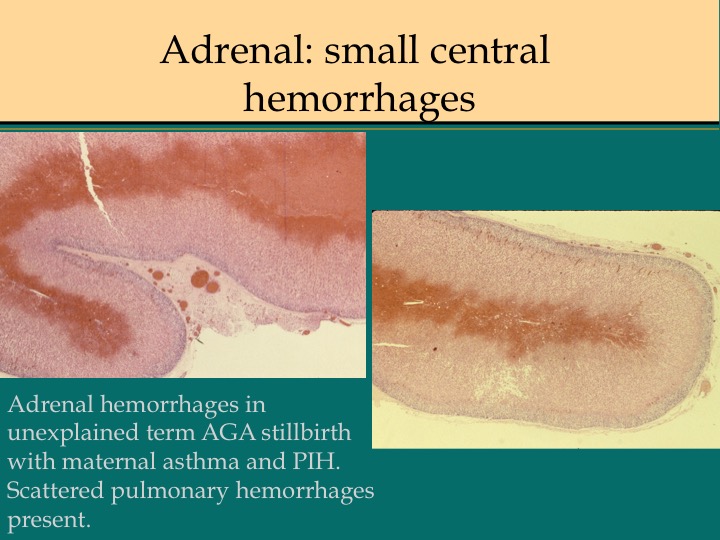
Before the advent of corrective surgery these infants died of cardiogenic shock in response to closure of the ductus arteriosus. Focal infarction an can also occur. Often in the adrenal the line between marked sinusoidal congestion and hemorrhage is hard to distinguish. The hemorrhage usually begins to obliterate the sinusoidal outline starting in the deepest fetal cortex. (Fig 10,11)
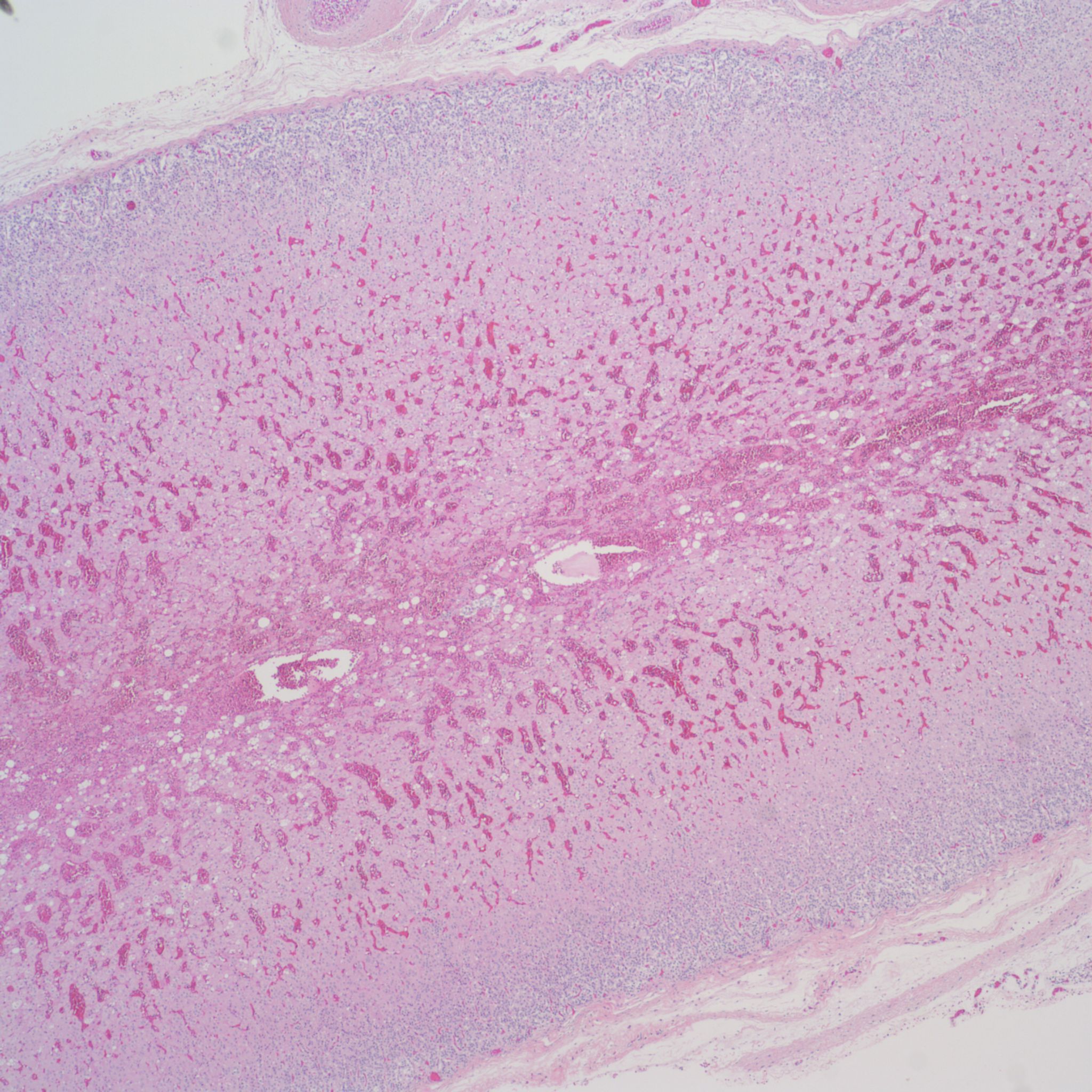
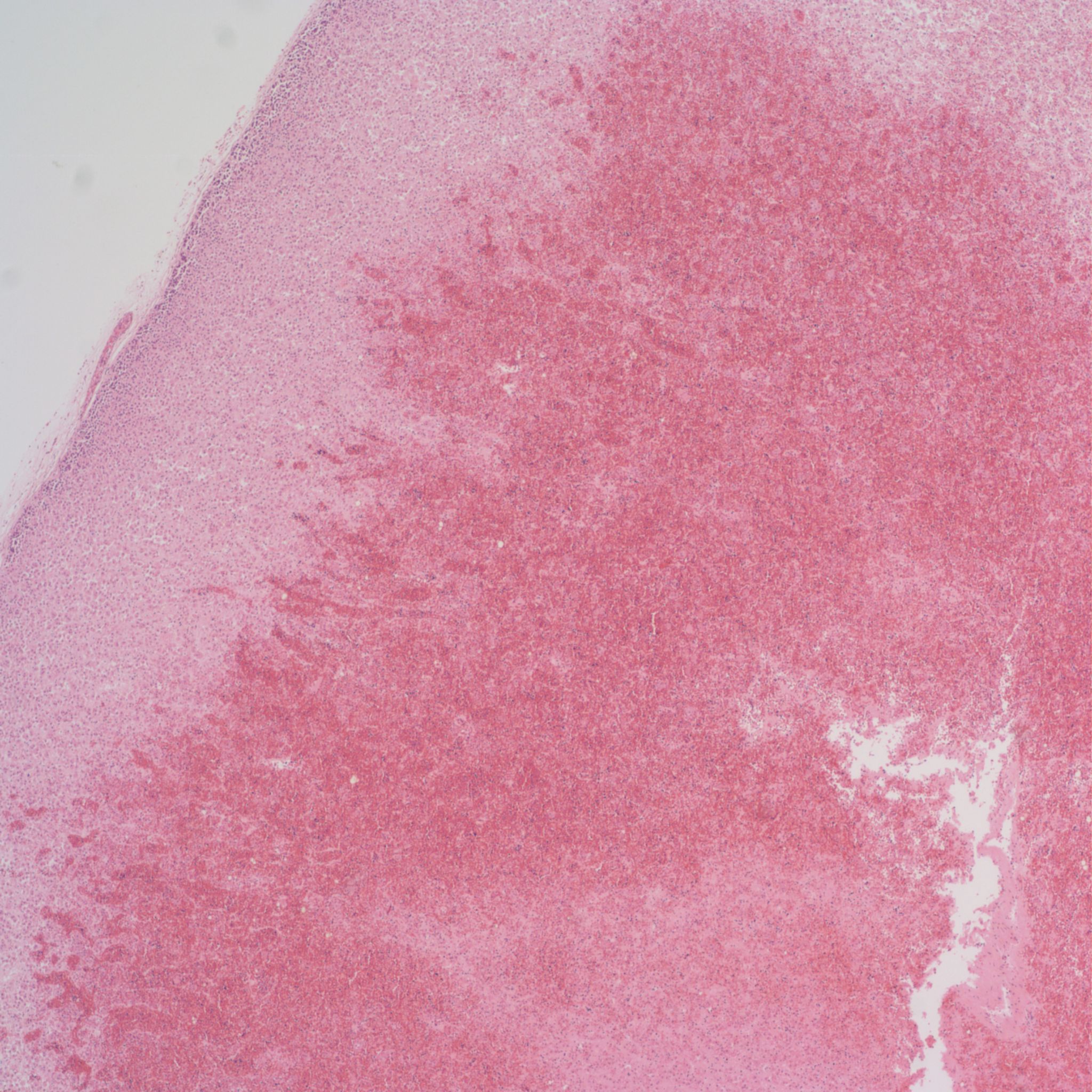
In the context of fetal infection, I accept adrenal hemorrhage as evidence of fetal septic shock similar to W-F hemorrhages, however, but I am unaware of evidence for this. A study by Jacques and Qureshi found that adrenal hemorrhage correlated with other evidence of acute asphyxia such as abruption, intrathoracic petechiae and lack of thymic involution[11]. Figs (12-14)
Fig 12-14: These are slides from a case conference. The first slide demonstrates the adrenal with hemorrhage. The next slide demonstrates the lung with extensive congenital pneumonia with overgrowth of pale basophilic cocci. The third slide demonstrates on the left a cluster of gram + cocci on the fetal membranes and a sparse maternal neutrophil infiltration at the junction of the chorionic epithelium and connective tissue (arrow) (Brown Gram). The right panel shows a similar cluster of cocci on the surface of the umbilical cord (H&E). The lung culture grew Group B beta hemolytic Streptococcus.


The role of the fetal cortex of the adrenal:
The role fetal adrenal cortex is a primate organ, not present in other animals. Liggins and colleague’s discovery is well known: the simple relationship of a maturing hypothalamic-pituitary-adrenal axis increases fetal cortisol which results in maturation of fetal organs and increases the estrogen to progesterone blood ratio that in turn leads to labor, at least in sheep. The problem is that this simple and elegant paradigm does not hold in humans because the critical enzyme CYP 17 that is upregulated by cortisol that in turn decreases the conversion of pregnenolone to estradiol in the sheep is absent in humans. One theory is that humans need to delivery immature infants because if the brain developed fully, the head could not be delivered through the pelvis. The primate fetal cortex could be a link in this delayed labor. If so, can the autopsy contribute to any understanding of the initiation of labor? Is there a volume change with labor or a detectable upregulation in some other adrenal enzyme? Is it even possible that there are detectable adrenal changes in the onset of preterm labor that might differ from induced preterm delivery? One problem is that we are looking at disease that has resulted in fetal death whether simple previable delivery or delivery induced due to fetal stress or maternal preeclampsia. Another is that proteomics, or even identifying more specific markers, are hampered by postmortem lysosomal degradation. Except for identifying pesticides in preterm labor[29], there does not appear to be published literature on cord blood composition and preterm labor, or for that matter induced and spontaneous term labor.
There have been multiple studies demonstrating the lack of a fetal cortex in anencephalic fetuses and two authors with pathological correlation have demonstrated a tendency to prolonged gestation in fetuses with loss of the pituitary adrenal axis and small fetal cortex[19, 20, 30]. In one very careful study which separated out complicated cases for example with polyhydramnios from normal deliveries of liveborn anencephalic infants with autopsy proven small adrenals the authors found an approximately 2 week delay in delivery compared to controls[30]. However, adjusting for the loss of brain weight, the infants were delivered at the normal weight and placental to fetal weight ratios. Thus, the delay could have been related to delayed growth and maturation, or to the absent fetal adrenal.
Fetal monkey adrenalectomy resulted in 6 term labors, and 2 late labors. Histology demonstrated that some adrenal rests remained[31].
Cortisol measurements in umbilical cord serum have consistently shown increases with gestational age and an abrupt increase with labor[32]. Cortisol is needed to stimulate the maturation of the lung, as demonstrated in clinical practice by the administration of betamethasone, a synthetic hormone that does cross the placental barrier, to induce lung maturation in preterm infants. How can cortisol be increased without effecting the amount of DHEA produced. In vitro cultured baboon fetal adrenal cells produced DHEA in response to ACTH and prolactin, but estradiol inhibited the effect of ACTH on DHEA production. This could be a mechanism to increase cortisol without affecting the DHEA estrogen system[33]. This must be only partially effective if it occurs, as the levels of DHEA increase with gestation in human gestation[34]. The human serum umbilical levels of cortisol increase with increasing gestation with a marked increase at term, and with a consistently higher level with labor independent of gestation based on 156 deliveries[35]. In chronically catheterized fetal monkeys, corticosteroid is markedly increased with spontaneous labor[36].
Bottom line:
The adrenal functions as a stress organ in utero. The accumulation of lipid is likely a response to ACTH but the specifics of its control and any advantage to the fetus such as the accelerated involution of the thymus is unclear. The mechanism and function of congestion and hemorrhage with shock needs further elucidation. The meaning of adrenal cytomegaly needs further clarification but is a potential marker of Beckwith Wiedemann syndrome. The adrenal is a major target for foci of necrosis with viral inclusions in congenital herpes infection. The pathologist can document organ weight, lipid accumulation, hemorrhage, benign focal cytomegaly and viral infection.
References:
6. Phillippe, M., Fetal catecholamines. Am J Obstet Gynecol, 1983. 146(7): p. 840-55.
24. Kalousek, D.K., Case 6. Adrenal hypoplasia in triploidy. Pediatr Pathol, 1984. 2(3): p. 359-62.
